Avery Dutcher, D.O., Oklahoma State University Center for Health Sciences, Tulsa OK
Nicole Cueli, D.O., Oklahoma State University Center for Health Sciences, Tulsa OK
Liming Fan, M.S., Oklahoma State University Center for Health Sciences, Tulsa OK
Al Rouch, Ph.D., Oklahoma State University Center for Health Sciences, Tulsa OK
Partially funded by the National Institute of General Medical Sciences of the National Institutes of Health through Grant Number 8P20GM103447
Abstract
Background: Research demonstrates that important sex differences exist in dietary-induced increase in blood pressure. The objective of this study was to investigate sex differences in blood pressure and renal handling of sodium in mice consuming a high-salt and high-fructose (HSHF) diet.
Methods: Healthy 4-week-old male and female mice (n=6/group) were placed in metabolic cages for six weeks. After the baseline week all mice consumed the HSHF diet consisting of 4% salt chow with a drinking solution of 20% fructose and 1% NaCl for 4 weeks. This was followed by a recovery week with mice on the normal diet with water. Blood pressure was measured daily via the tail cuff technique. Daily measurements of sodium intake and output were measured. Real-time PCR was conducted to measure relative expression of key renal sodium transporters.
Results: Systolic blood pressure (SBP) significantly increased in male and female mice during the HSHF period. SBP decreased in male mice in the recovery week but remained elevated in female mice. Female mice showed higher sodium retention during the HSHF period via the Nae/Nai ratio (62±5% vs 75±5%, p<0.01). Molecular expression of renal sodium transporters was significantly higher in the female mice.
Conclusion: We conclude that female mice are not protected from the HSHF dietary-induced increase in blood pressure. Higher sodium retention might explain the lack of protection in female mice. This study challenges the current position that females possess protective mechanisms against dietary induced increase in blood pressure, at least with high fructose.
Keywords: Blood Pressure, Diet, Sex Difference, Fructose. Salt
Introduction
The addition of high fructose corn syrup to food products and particularly to soft drinks in the 1970s has led to high levels of fructose consumption, and can be held responsible for contributing to the obesity epidemic 1,2. The link between high salt consumption and hypertension has been known for many years 3,4. Dietary consumption of both high fructose and high salt enhance the health problems associated with consumption of either salt or fructose alone.
Sex differences have been reported regarding the effects of high fructose consumption 5,6. Sex differences have also been shown with respect to blood pressure regulation, cardiovascular, and renal function 7-11. The impact of fructose on the kidney has important consequences to the renal handling of salt and water and thus to overall sodium balance. By researching how high fructose and salt consumption affects the renal handling of sodium, blood pressure, and various pathologies, newer and more effective therapies for metabolic syndrome and hypertension can be developed.
The objective of this research study was to investigate sex differences in blood pressure regulation and renal handling of sodium in mice consuming a high salt, high fructose (HSHF) diet. Since females are often protected from causes of hypertension compared to males, an important question for this research is to determine if females are protected from high blood pressure when consuming high fructose and salt. Other objectives included how a HSHF diet affects the renal handling of sodium and sodium balance in male and female mice, and if sex differences exist with respect to the molecular expression of key sodium transporters in the kidney.
Methods
Animals: Male and female CD-1, 4-wk-old mice were purchased from Envigo Laboratories (Indianapolis, IN). Upon arriving to the OSU-CHS animal facilities, mice were placed in plastic bins and consumed normal diet with water before the start of the study. All mice were pathogen free and healthy. The OSU-CHS IACUC approved all experimental procedures.
Procedures: Mice (n=6/group) were placed individually in metabolic cages for 5 days/week throughout the study. On weekends, these mice were placed individually in a plastic bin but maintained the specific diet assigned to the weekly period. Separate mice used for the molecular experiments (see below) remained in plastic bins and on the appropriately assigned diet. Mice consumed normal diet with water in the first or baseline week. All mice were then placed on the HSHF diet consisting of 4% salt chow (Teklad, TD 120254) with a drinking solution of 20% fructose and 1% NaCl for the next 30 days. This was followed by a 9-day recovery period with all mice again consuming normal diet with water.
Blood Pressure Measurement: We performed the tail-cuff technique to measure systolic blood pressure (SBP) using the CODA device from Kent Scientific (Torrington, CT) 12. Briefly, this technique is performed with the mouse in a restrainer where the tail is exposed and placed in an occlusion cuff. Basically, the tail-cuff technique measures blood pressure via sphygmomanometry. This non-invasive technique provides accurate blood pressure measurement and is widely used 13-16. The primary concern of this technique is the stress level of the mouse in the restrainer. Two or three sessions of about 15 – 20 minutes/session in the restrainer adequately prepare a mouse for the technique. Mice in this study were give one or two sessions per day for three days prior to blood pressure recording. Measurements were taken on each mouse assigned to the metabolic cages 4-5 days/week and systolic blood pressure (SBP) measurements averaged weekly for females and males are reported in the Results.
Physiological Measurements: Daily measurements of mice in metabolic cages included body weight, food & water (or fluid) intake, urine analysis for urine flow rate (UFR) and Na+ concentration (Una) which was measured using an EasyLyte® electrolyte analyzer. Sodium excretion (Nae) was determined daily via: UNa (mEq/L) x UFR (ml/day). Sodium balance = Sodium Input (calculated from food and fluid intake) minus Sodium Output (calculated from Nae).
Molecular Experiments: Quantitative real-time PCR (qt-PCR) was employed to measure relative expression of key sodium transporters in the kidney. These studies were conducted at the end of the baseline period, at the end of the first two weeks and last two weeks of the HSHF period, and at the end of recovery period. Mice were anesthetized and killed via cervical dislocation followed by removal of the kidneys. Total RNA was extracted from renal cortical tissue using the MELTTM Nucleotide Isolation kit (Ambion) and cDNA was synthesized using the RT2 first-strand kit (SA Biosciences) and tested for quantity and quality via nano-drop technology using an Implen® (N60) nanophotometer. Real-time PCR was conducted using custom-made PCR arrays (96-well plates) made by Qiagen SA Biosciences and designed with specific primers for the following mouse Na+ transporters: 1) ENaCᾳ (Scnn1a); 2) ENaC (Scnn1b); 3) ENaCγ (Scnn1g); 4) NKCC (Slcl2a1); 5) NCC (Slc12a3); 6) NHE3 (Slc9a3); 7) NHE2 (S19a2); 8) NaPi2A (Slc34a1); 9) SGLT (Slc5a12); 10) ᾳNaK ATPase (Atp1a1); 11) ATR1 (Agtra1a); and 12) ATR2 (Agtr2). Primers were tested and demonstrated 95-100% amplification efficiency using the PCR reagents provided by SA Biosciences.
Quantitative-PCR was performed via Sybr-green technology on an Opticon 2 (MJ Research) thermocycler. The PCR arrays contained primers for the following reference genes: 1) beta actin (Actβ); 2) glyceraldehyde-3-phosphate dehydrogenase (Gapdh); 3) hypoxanthineguanine phosphoribosyl transferase (Hprt); 4) glucuronidase beta (Gusb); 5) heat shock protein 90 alpha (cytosolic) class B member (Hsp90ab1). Qiagen SA Biosciences provides software for analysis via Student t-test of relative expression via the ΔΔCt method 17. Male vs female comparisons are analyzed in triplicate and overall, minimal criteria for PCR analysis were met 18.
Statistics: Data are shown as mean ± sem. Repeated measures 2-way ANOVA was used to analyze SBP, Nae, and sodium balance. Student unpaired t-test was used for simple comparisons of sodium retention, sodium balance, and sodium excretion. Molecular expression data were analyzed via t-test and expressed as fold difference, female-to-male. Prism 9 (GraphPad Software, version 9.3.1) was used for all statistical analysis. Differences of P < 0.05 were considered statistically significant.
Results
Figure 1 shows the SBP of male and female mice throughout the study. SBP in the baseline week was not different between males and females and SBP did not increase after the first week on HSHF consumption. SBP increased in both male and female mice in week 3 and remained elevated throughout the HSHF period. Male SBP decreased in male mice during recovery but not in female mice.
Figure 1. Systolic Blood Pressure
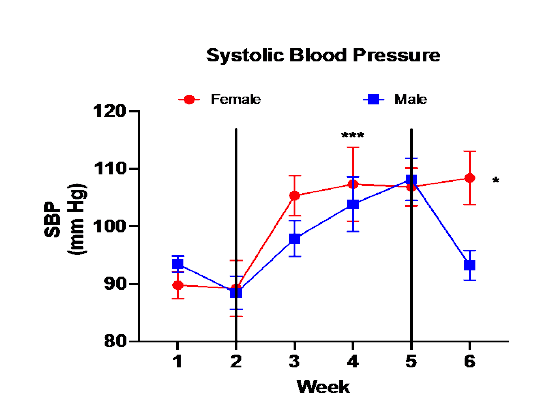
Figure 1. SBP (mean ± sem) of male and female mice during baseline week (1) and the four weeks with mice on HSHF diet (2, 3, 4, 5), and the recovery period where mice consumed normal diet with water (6). SBP was measured daily of each mouse assigned to the metabolic cages and values were averaged for the specific week. SBP did not increase after the first week on the HSHF diet (2). (*** - SBP in weeks 3, 4, & 5 > SBP in baseline week (1) and week 2, p<0.001 via 2-way repeated-measures ANOVA). SBP in male mice decreased in the recovery period in male mice but not in female mice. (* - female SBP > male SBP in recovery period p<0.01).
Figure 2 shows sodium excretion and sodium balance data. Measurements were taken daily while mice were in metabolic cages. Female mice tended to have lower Nae and higher sodium balance compared to male mice.
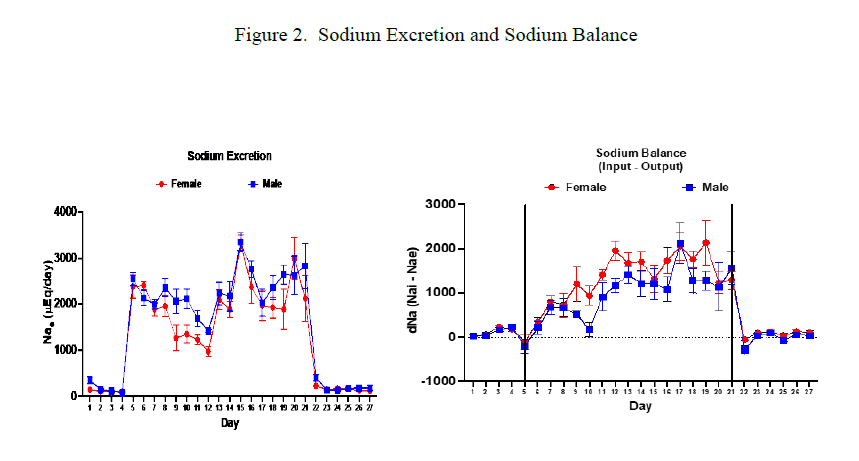
Figure 2. Sodium excretion (mean ± sem) data shown in the graph on the left. Baseline and recovery periods clearly shown by the low Nae values on either side. Repeated measures ANOVA resulted in no significant difference between male and female mice. Female mice Nae was non-significantly lower than Nae for male mice during the HSHF consumption period. Graph on right shows daily sodium balance data (mean ± sem). Repeated measures ANOVA showed no significant difference between male and female mice, however female values were non-significantly higher.
Figure 3 shows the measurement of Nae-to-Nai ratio during the HSHF period in male and female mice. Nae/Nai for female mice is significantly lower for female mice which suggests higher sodium retention for female.
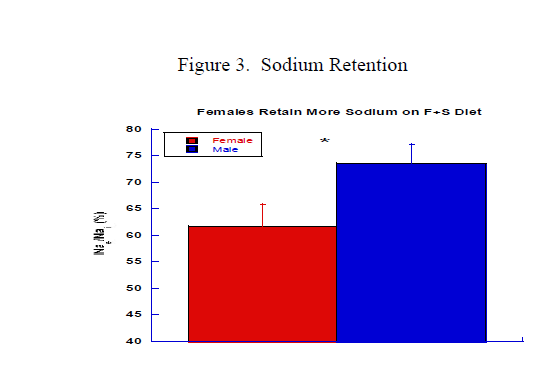
Figure 3. Graph shows average ratio of the ratio of sodium excretion-to-sodium intake during the HSHF consumption period. Females had significantly lower ratio than males (62.0 ± 4.4 vs 74.8 ± 3.7, * p<0.05). Data indicate that females retained more sodium throughout the HSHF period.
Moreover, during the recovery period females also had lower ratio compared to males (58.4 ± 4.3 vs 83.2 ± 6.0, p<0.01) which could explain the SBP results in the recovery period.
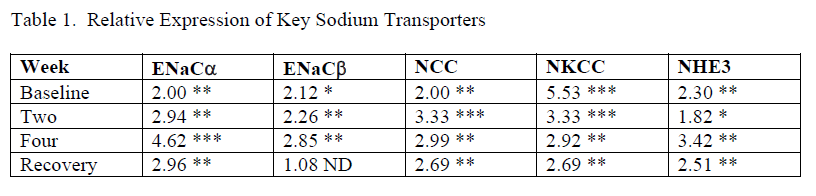
Table 1. Data showing relative expression with values of fold difference (FD) between female and male, i.e, male used as reference. All FD values in the table indicate higher expression in female kidneys. Results from baseline, week two, and week four come from the separate mice placed in bins (see Procedures in Methods), n=4/group. Results from the Recovery week come mice used in metabolic cages, n=6/group. One FD value in the study not shown in this table resulted in a significantly higher expression in male kidney – NaPi, sodium-phosphate cotransporter in proximal tubule, p<0.01). * p<0.5, ** p<0.01, *** p<0.001
Discussion
The obesity epidemic and high prevalence of hypertension have become global health concerns challenging the medical profession and placing enormous financial costs for treatment of the associated pathologies. The high risks associated with obesity and hypertension are modifiable with proper diet and exercise. While we strive to convince the public to engage in healthy habits, we should expect to continue treating patients with obesity and hypertension for years to come.
HSHF consumption in society contributes to both obesity and hypertension and research into the pathophysiology of HSHF in animals and humans has been quite extensive for the past several years. We set out to add to this body of knowledge by studying sex differences in mice consuming a HSHF diet. Our major goal was to determine if HSHF diet increased blood pressure in both male and female mice. Research has demonstrated that females are protected against high salt-induced increase in blood pressure compared to males19,20. So, a major question posed in this study was: would females be protected from elevated blood pressure during the consumption a HSHF diet? Another important issue we wanted to investigate was the renal handling of sodium. Since the kidneys control blood pressure via adjusting the renal excretion of sodium, it becomes important to measure Nae and sodium balance in both sexes. Finally, we investigated the molecular expression of major sodium transporters to provide insight to the results on blood pressure and sodium balance.
Results showed that the HSHF diet raised SBP in both male and female mice although it took until the second week on the diet to raise SBP (Figure 1). Our findings demonstrated female mice had no protection against increased SBP. In fact, SBP appeared to increase faster in females with higher SBP compared to males in week two of HSHF consumption. Of particular importance is the observation that SBP decreased in male mice but not in female mice during the recovery period. Thus, the addition of fructose to the diet could be responsible for the loss of protection and this might be explained by the renal handling of sodium.
The results on sodium balance in this study basically showed that both males and females retained a substantial amount of sodium during the HSHF diet (Figure 2). Based on our study female mice do not excrete more sodium than males when consuming high salt. This suggests that this mechanism is not the reason for protection from high blood pressure under high salt conditions in females. Estrogen is believed to play a major role in protection against hypertension, but it should also be remembered that research from many years ago demonstrated the salt-retaining effects of estrogen 21-23. Since then, studies have examined the roles of sex steroids on kidney function clearly demonstrating that both estrogens and androgens have important consequences on many renal functions 7-9,11,24,25.
Most relevant to this study, is a report by Soleimani and Alborzi who proposed that high fructose intake changes renal transport activity increasing sodium and chloride reabsorption in the proximal tubule 26. These authors presented a model involving fructose-induced increase in activity of the membrane transporters Glut5 which transports fructose, the sodium-hydrogen exchanger NHE3, which transports sodium, and the putative anion transporter 1 (PAT1) which transports chloride. This model could explain how high dietary fructose increases salt retention and induces hypertension. Further research is required to confirm this mechanism.
Our study showed that both male and female mice retained sodium during the HSHF consumption and females probably retained more than males as shown in figures 2 & 3. This might explain the loss of protection against the HSHF-induced increase in SBP in female mice. The expression data in Table 1 shows higher expression of renal transporters in the distal nephron of female kidneys. Female kidneys expressed higher relative expression of key sodium transporters including NKCC, NCC, NHE3, and ENaC than male kidneys. This now appears to be a consistent finding with other studies we have conducted and is consistent with other research into the sexual dimorphism of renal transporters 27 28. This could also explain the high SBP during the recovery period in female mice compared to male mice.
Future studies looking at the role of estrogen are being planned to determine if its presence increases sodium transport within the kidney. The study will repeat these experimental procedures in ovariectomized mice with and without estrogen supplementation. If estrogen is responsible for higher expression, the estrogen-supplemented mice should show higher expression of sodium transporters and different blood pressure profile than the placebo-treated mice.
Conclusion
HSHF consumption increased SBP in male and female mice. SBP decreased in male mice but not in female mice during the recovery period. We conclude that female mice are not protected from HSHF-induced increase in blood pressure. Both male and female mice retained sodium during the HSHF consumption period. Female mice had higher expression of some key renal sodium transporters, and this could be responsible for higher sodium retention and subsequent higher SBP in the recovery period. This study challenges the current position that females possess protective mechanisms against dietary-induced increases in blood pressure.
REFERENCES
1. Gordish KL, Kassem KM, Ortiz PA, Beierwaltes WH. Moderate (20%) fructose-enriched diet stimulates salt-sensitive hypertension with increased salt retention and decreased renal nitric oxide. Physiological Reports. 2017;5(7):e13162.
2. Marriott BP, Cole N, Lee E. National estimates of dietary fructose intake increased from 1977 to 2004 in the United States. J Nutr. 2009;139(6):1228S-1235S.
3. Elijovich F, Weinberger M, Anderson CAM, et al. Salt Sensitivity of Blood Pressure: A Scientific Statement from the American Heart Association. Hypertension. 2016;68:e7-e46. doi:10.1161/HYP.0000000000000047
4. Takahashi H, Yoshika M, Komiyama Y, Nishimura M. The central mechanism underlying hypertension: a review of the roles of sodium ions, epithelial sodium channels, the renin-angiotensin-aldosterone system, oxidative stress endogenous digitalis in teh brain. Hyper Res. 2011;34:1147-1160.
5. Galipeau D, Verma S, McNeill JH. Female rats are protected against fructose-induced changes in metabolism and blood pressure. Am J Physiol Heart Circ Physiol. 2002;283:H2478-H2482. doi:10.1152/ajpheart.00243.2002
6. Sharma N, Li L, Ecelbarger CM. Sex differences in renal and metabolic responses to a high-fructose diet in mice. Am J Physiol Renal Physiol. Mar 1 2015;308(5):F400-10. doi:10.1152/ajprenal.00403.2014
7. Iliescu R, Reckelhoff JF. Sex and the kidney. Hypertension. 2008;51:1000-1001.
8. Pechère-Bertschi A, Burnier M. Female sex hormones, salt, and blood pressure regulation. Am J Hyper. 2004;17:994-1001.
9. Pechère-Bertschi A, Burnier M. Gonadal steriods, salt-sensitivity and renal function. Curr Opin Nephrol Hyper. 2007;16(1):16-21.
10. Reckelhoff JF. Gender differences in the regulation of blood pressure. Hypertension. 2001;37(5):1199-1208.
11. Reckelhoff JF. Sex and sex steroids in cardiovascular-renal physiology and pathophysiology. Gender Medicine. 2008;5(Suppl. A):S1-S2.
12. Feng M, Whitesall S, Zhang Y, Beibel M, D'Alecy L, Dipetrillo K. Validation of volume-pressure recording tail-duff blood pressure measurements. Am J Hyper. 2008;doi 10.1038/ajh.301:301-310.
13. Aljunaidy MM, Morton JS, Cooke CL, Davidge ST. Maternal vascular responses to hypoxia in a rat model of intrauterine growth restriction. Am J Physiol Regul Integr Comp Physiol. Dec 1 2016;311(6):R1068-R1075. doi:10.1152/ajpregu.00119.2016
14. Mishra JS, Hankins GD, Kumar S. Testosterone downregulates angiotensin II type-2 receptor via androgen receptor-mediated ERK1/2 MAP kinase pathway in rat aorta. J Renin Angiotensin Aldosterone Syst. Oct 2016;17(4)doi:10.1177/1470320316674875
15. Riera M, Anguiano L, Clotet S, et al. Paricalcitol modulates ACE2 shedding and renal ADAM17 in NOD mice beyond proteinuria. Am J Physiol Renal Physiol. Mar 15 2016;310(6):F534-46. doi:10.1152/ajprenal.00082.2015
16. Wang H, Sun X, Chou J, et al. Cardiomyocyte-specific deletion of the G protein-coupled estrogen receptor (GPER) leads to left ventricular dysfunction and adverse remodeling: A sex-specific gene profiling analysis. Biochim Biophys Acta. Aug 2017;1863(8):1870-1882. doi:10.1016/j.bbadis.2016.10.003
17. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT method. Methods. 2001;25:402-408.
18. Bustin SA, Benes V, Garson JA, et al. The MIQE guidelines: Minimal Information for Publication of quantitative real-time PCR experiments. Clin Chem. 2009;55(4):611-622.
19. Brinson KN, Rafikova O, Sullivan JC. Female sex hormones protect against salt-sensitive hypertension but not essential hypertension. Am J Physiol Regul Integr Comp Physiol. 2014;307:R149-R157.
20. Xue B, Pamidimukkala J, Hay M. Sex differences in the development of angiotensin II-induced hypertension in conscious mice. Am J Physiol Heart Circ Physiol. 2005;288:H2177-H2184.
21. Preedy JRK, Aitken EH. The effect of estrogen on water and electrolyte metabolism. III. Cardiac and renal disease. J Clin Invest. 1956;35:443-451.
22. Preedy JRK, Aitken EH. The effect of estrogen on water and electrolyte metabolism. II Hepatic disease. J Clin Invest. 1956;35:430-442.
23. Thorn GW, Engel LL. The effect of sex hormones on the renal excretion of electrolytes. J Exper Med. 1938;68:299-305.
24. Cheema MU, Irsik DL, Wang Y, et al. Estradiol regulates AQP2 expression in the collecting duct: a novel inhibitory role for estrogen receptor alpha. Am J Physiol Renal Physiol. Aug 15 2015;309(4):F305-17. doi:10.1152/ajprenal.00685.2014
25. Hutchens MP, Nakano T, Kosaka Y, et al. Estrogen is renoprotective via a nonreceptor-dependent mechanism after cardiac arrest in vivo. Anesthesiology. 2010;112(2):305-405. doi:10.1097/ALN.0b013e3181c98da9
26. Soleimani M, Alborzi P. The role of salt in the pathogenesis of fructose-induced hypertension. Int J Nephrol. 2011;2011:392708. doi:10.4061/2011/392708
27. Veiras LC, Girardi ACC, Curry J, et al. Sexual Dimorphic Pattern of Renal Transporters and Electrolyte Homeostasis. J Am Soc Nephrol. Dec 2017;28(12):3504-3517. doi:10.1681/ASN.2017030295
28. Li J, Hatano R, Xu S, et al. Gender difference in kidney electrolyte transport. I. Role of AT1a receptor in thiazide-sensitive Na(+)-Cl(-) cotransporter activity and expression in male and female mice. Am J Physiol Renal Physiol. Aug 1 2017;313(2):F505-F513. doi:10.1152/ajprenal.00087.2017